The LabCoat Guide to Pesticide Mode of Action: Insecticides – Part I
In this third article in the LabCoat Guide to Pesticide Mode of Action, I discuss conventional Insecticides and their Biochemical Mode of Action.
A little about my background
I am a plant scientist with a background in Molecular Plant Biology and Crop Protection. 20 years ago, I worked at Copenhagen University on photosynthetic responses to stress in crops. Subsequently, I worked in Australia on molecular defense mechanisms induced by Phylloxera attack in grapevine.
At that time, biology-based crop protection strategies had not taken off commercially, so I transitioned to conventional (chemical) crop protection R&D at Cheminova, later FMC.
During this period, public opinion, as well as increasing regulatory requirements, gradually closed the door of opportunity for conventional crop protection strategies, while the biological crop protection technology I had contributed to earlier began to reach commercial viability.
From January 2018, I will consolidate 17+ years of industry experience in BioScience R&D management with my academic research background, to provide independent Strategic R&D Management as well as Scientific Development and Regulatory support to AgChem & BioScience organizations developing science-based products.
For more information, visit BIOSCIENCE SOLUTIONS – Strategic R&D Management Consultancy.
***
Insecticides are substances toxic to insects that are used to destroy unwanted pests, as well as their eggs and larvae. Insecticides control pests by interfering with their growth or reproduction, or by repelling insects from treated crops. This is achieved through a number of different Modes of Action (MOA).
Modes of Action of Insecticides
MOA may be differentiated into Mechanism of Action, referring to the biochemical site of herbicide activity, and Mode of Action – the associated symptoms that occur following insecticide application.
In the following, Mode of Action will be used to describe both the site of action as well as associated insect symptoms. The chemical structure of the insecticide active ingredient (AI) usually defines its mode of action by determining its uptake and systemicity, as well as its ability to reach and bind to the target site – the physical location where are the insecticide acts.
A sound knowledge of insecticide mode of action is important for determining how to optimally use an insecticide, as well as for understanding the underlying mechanisms of resistance, selectivity and toxicity to beneficial insect species, fish, birds and mammals.
In crop protection R&D, elucidating insecticide mode of action requires years of research, coupling biochemical and molecular biological knowledge with insect physiology and the timing and nature of symptom development.
Insecticides may target insects through a number of modes of action. They may target insects by inhibiting energy production, typically by inhibiting mitochondrial respiration, while insecticides such as chitin synthesis inhibitors (CSIs) target the production of insect exoskeletons, and prevent insect moulting.
Inhibitors of the endocrine system or insect growth regulators (IGR) act on the hormonal system of insects, preventing them from maturing and reproducing. Insecticides affecting the water balance of insects (desiccants) break down the waxy coating of their exoskeletons, resulting in the water loss and death.
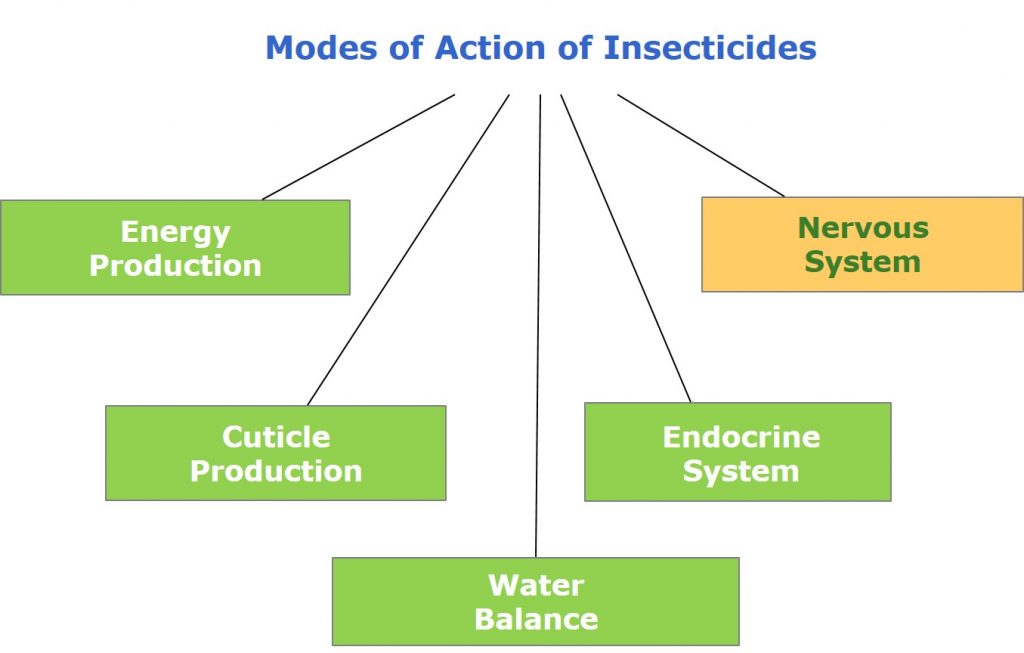
Figure 1: Modes of action of insecticides.
However, most commercial insecticides target the nervous system of insects.
Insecticides targeting nervous systems
Most insecticides targeting nervous systems may be grouped into those inhibiting the normal function of Cholinergic and GABA synapses, respectively. Within these groupings, insecticides target specific neural functions, such as the uptake of ions across neural membranes, or by inhibiting specific enzyme functions or by preventing the binding or release of neurotransmitters from their respective receptors.
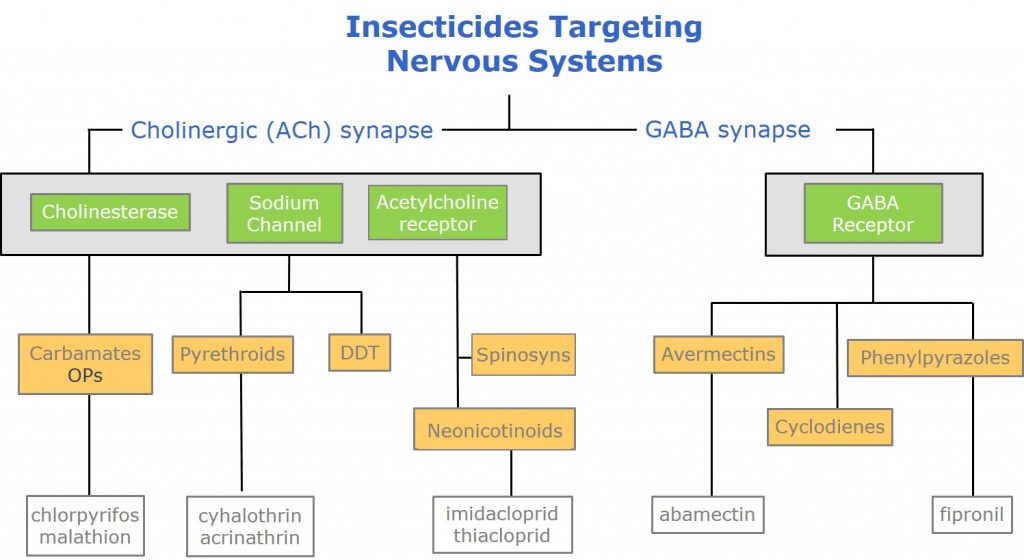
Figure 2: Insecticides targeting nervous systems.
Nervous systems in insects
To understand the mode of action of insecticides targeting nervous systems, it is necessary to consider how the nervous system functions. The nervous system of an insect can be divided into a “brain” (comprising three pairs of ganglia, or nerve cell bundles) which are connected to a ventral nerve cord made up of interconnected nerve cells (neurons) along which electrical charges – called impulses – travel.
These impulses are driven by the so-called action potential – a rapidly changing spike of depolarisation and repolarisation induced by the activation of ion channels in the nerve cell membrane. It is through this action potential that messages are sent from external sensors to the brain and back.
Neurons may be divided into three categories, depending on their function. Sensory neurons carry messages from “sense” receptors toward the insects “brain”, while motor neurons stimulate responses in muscles and glands. Association neurons transmit signals within the insect’s nervous system.
In insects, disruption of the nervous system by insecticides will result in paralysis or uncontrolled tremors, and ultimately death.
Individual nerve cells consist of two extremities, respectively termed dendrites and axon terminals. Between the axon terminal of the preceding nerve cell (presynaptic neuron) and the dendrites of the succeeding nerve cell (postsynaptic neuron), a microscopic junction or synapse is found.
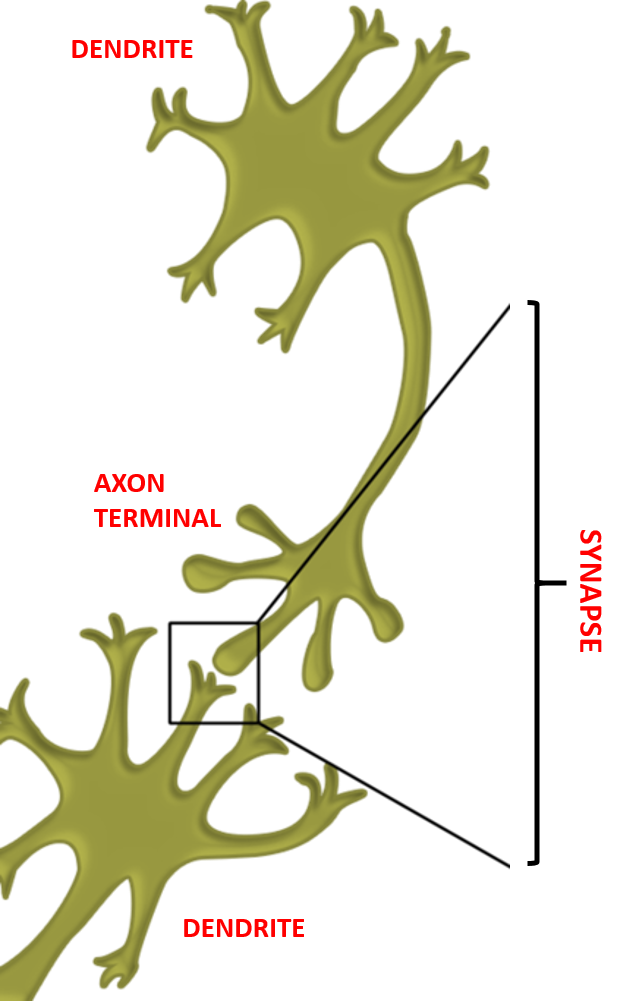
Figure 3: Insect nerve cells and synapse
Action potentials are transmitted from the dendrites to the axon terminal, at which stage the impulse must be converted from an electrical potential to a chemical signal capable of traversing the junction. This chemical signal or molecule is termed a neurotransmitter. Important neurotransmitters include Acetylcholine (ACh) and Gamma Amino Butyric Acid (GABA).
Inhibition of the Cholinergic (ACh) synapse by insecticides
The cholinergic synapse uses acetylcholine (ACh) as a neurotransmitter to transfer the action potential from the axon terminal of the presynaptic neuron to the dendrite of the postsynaptic neuron. These are termed activatory synapses, as they activate the subsequent neuron. In contrast, other synapses such as the GABA synapse are termed inhibitory synapses, as they reduce or inhibit post-synaptic neural activation.
In cholinergic neurons, nerve impulses arriving at the axon terminal trigger the release of ACh from neural vesicles. These neurotransmitter molecules are released from the axon terminal and traverse the synaptic cleft before binding to specific receptors on the surface of postsynaptic dendrites.
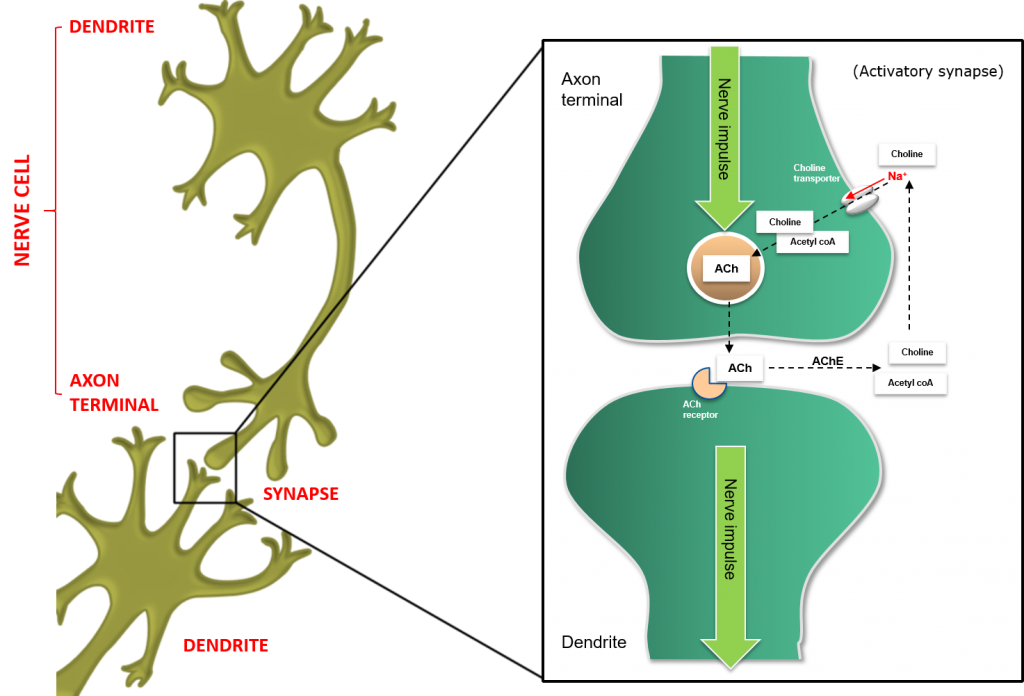
Figure 4: Function of the Acetylcholine (ACh) or Cholinergic synapse in insects.
Binding of ACh triggers an action potential, and the new nerve impulse is transmitted along the neuron. The ACh molecule is subsequently released from the receptor to which it was bound (ending nerve stimulation) and is degraded to its constituent components, Choline and Acetyl-CoA, through the actions of the enzyme Acetlycholine Esterase (AChE)
These building blocks are reabsorbed into the presynaptic neuron via voltage-gated sodium (Na+) channels in the neuronal membrane. Once reabsorbed, these building blocks are used to regenerate ACh neurotransmitters.
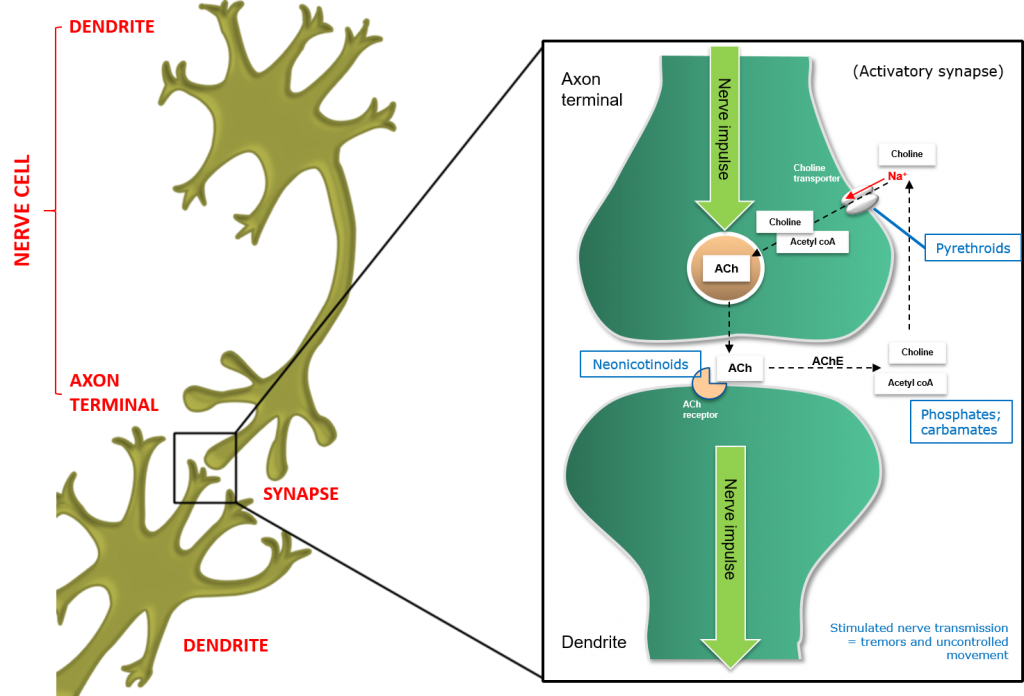
Figure 5: Inhibition of the Acetylcholine (ACh) or Cholinergic synapse in insects.
Neonicotinoid insecticides are synthetic analogs of the natural insecticide nicotine. Neonicotinoids compete with ACh neurotransmitters and competitively bind to the ACh receptor on the postsynaptic nerve, resulting in uncontrolled nerve stimulation (hyperstimulation), giving rise to symptoms of hyperactivity and tremors.
Organophosphate and Carbamate insecticides act by inhibiting AChE, preventing it from removing ACh from its receptor site. In the absence of active AChE, a stimulated nerve cannot return to its resting state, and the result is neural overstimulation and insect death.
Pyrethroid insecticides disrupt normal nerve function by binding to the voltage-gated sodium channels, preventing them from closing normally. This, in turn, gives rise to neural hyperactivity and insect death.
In the next article in this series, I discuss additional conventional Insecticide and BioInsecticide modes of action.
In other articles in this series, I focus on Herbicide, Fungicide and Insecticide modes of action, as well as basic Physicochemical and Formulation concepts related to Pesticide activity.
Harry Teicher is the founder of BIOSCIENCE SOLUTIONS and an Authorpreneur, providing organizations with Strategic- and Project Management as well as Development & Communication solutions. Follow him on Linkedin, Twitter and Facebook.